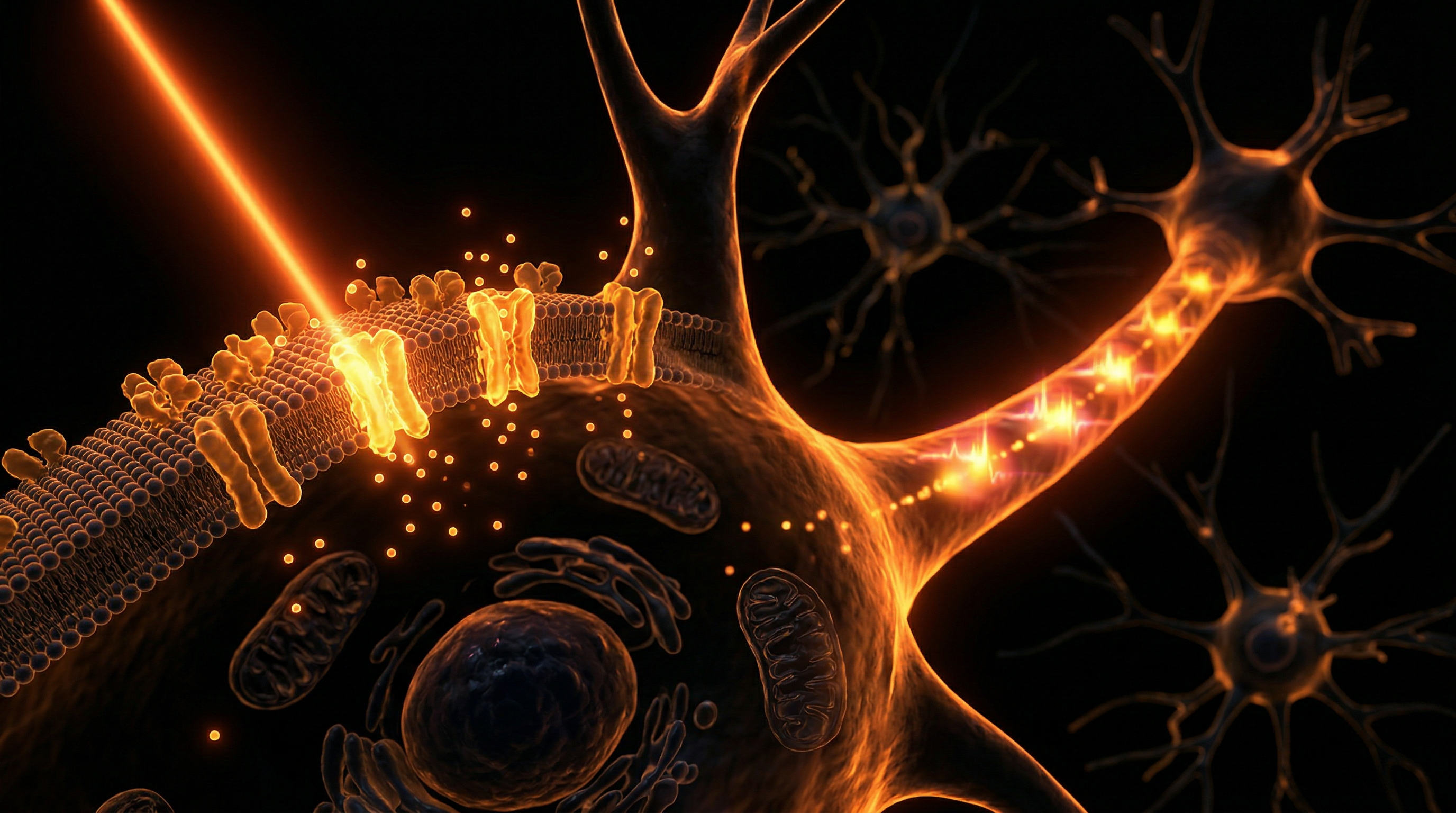
Optogenetics: Remote Control of Cellular Activity
Understanding the use of light-sensitive synthetic proteins to modulate heart rate and neuron firing. The article warns of the potential for non-invasive external manipulation of physiology.
Overview
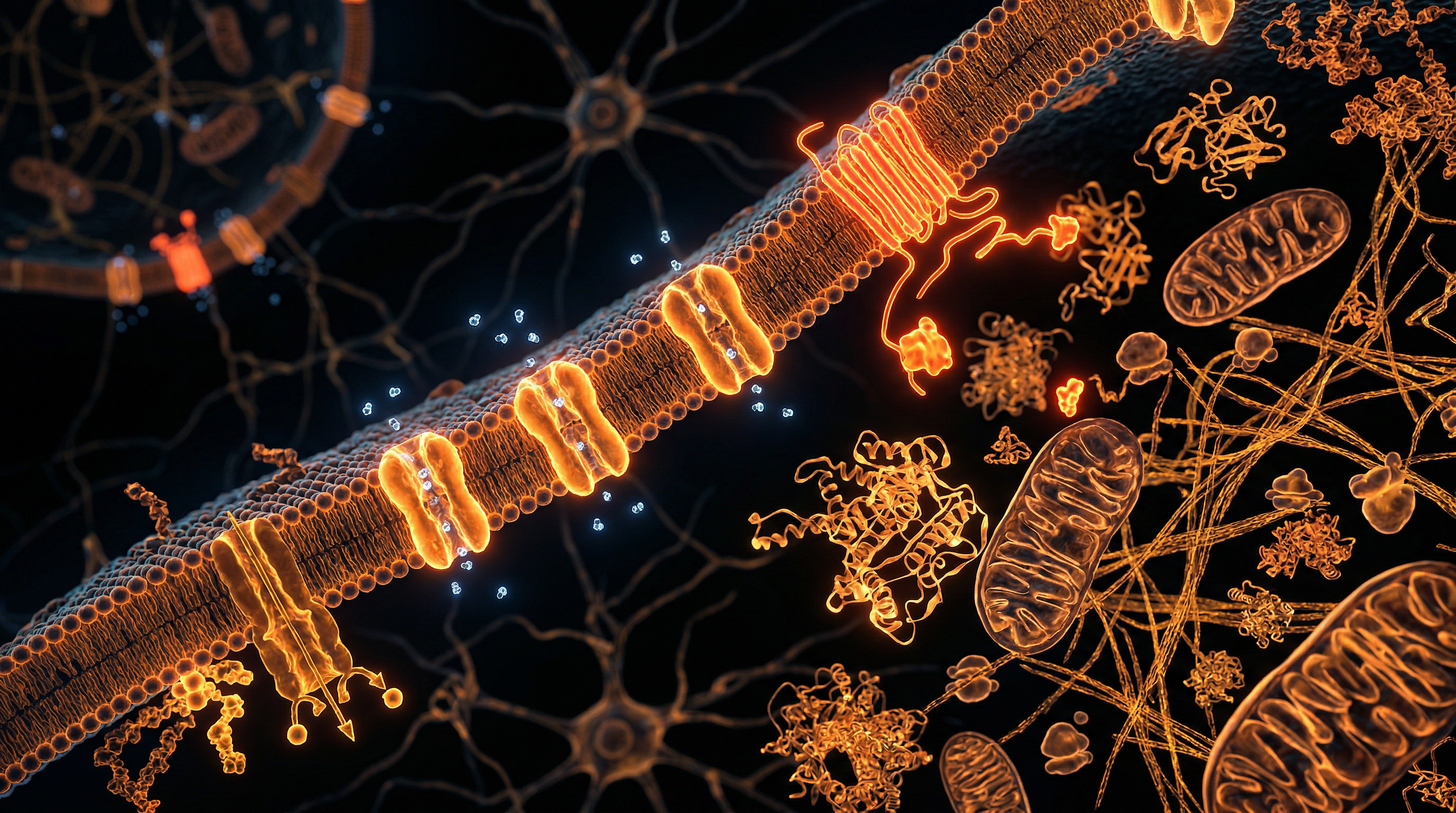
Optogenetics represents a seismic departure from the classical pharmacological paradigm, facilitating a "remote control" mechanism over cellular physiology with sub-millisecond precision. At INNERSTANDIN, we recognise this technology as the apex of modern synthetic biology—a sophisticated fusion of genetic engineering and photonic stimulation designed to override or augment endogenous bio-electrical pathways. Unlike systemic drug administration, which suffers from off-target effects and poor temporal resolution, optogenetics permits the interrogation of specific cellular circuits within complex biological matrices. The fundamental mechanism relies on the exogenous expression of microbial opsins—light-sensitive proteins—within defined cell populations. The most prominent of these, Channelrhodopsin-2 (ChR2), derived from the green alga *Chlamydomonas reinhardtii*, functions as a light-gated cation channel. Upon exposure to blue light (approximately 470 nm), the protein undergoes a conformational shift, allowing an influx of cations that depolarises the membrane and triggers immediate action potentials.
The systemic implications of this technology extend far beyond rudimentary neuroscientific inquiry. By utilising adeno-associated virus (AAV) vectors to deliver these opsin genes—a method refined through extensive research at UK institutions such as University College London and the University of Oxford—scientists can now modulate cardiac rhythm, pancreatic insulin secretion, and immune response with unprecedented accuracy. Peer-reviewed literature, including seminal studies published in *Nature* and *The Lancet*, highlights the transition of optogenetics from a laboratory tool to a clinical contender. For instance, the use of inhibitory opsins like Halorhodopsin (NpHR) or Archaerhodopsin (Arch) allows for the hyperpolarisation of cells, effectively "switching off" aberrant activity in conditions such as focal epilepsy or chronic neuropathic pain.
Furthermore, the integration of nanotechnology is pivotal; the development of upconverting nanoparticles (UCNPs) allows for the activation of deep-tissue opsins using near-infrared (NIR) light, which penetrates biological tissue more effectively than visible light, reducing the need for invasive fibre-optic implants. This level of control represents a radical shift in how we understand biological autonomy. At INNERSTANDIN, we assert that the "remote control" of cellular activity is not merely a therapeutic prospect but a fundamental re-engineering of the human biological interface. As we move toward the era of bio-electronic medicine, the ability to synchronise cellular firing patterns across systemic networks offers a high-density solution to currently intractable neurological and metabolic disorders, firmly establishing optogenetics as the cornerstone of 21st-century synthetic biology.
The Biology — How It Works
The operational core of optogenetics lies in the heterologous expression of microbial opsin genes—specifically those encoding light-activated ion channels, pumps, and receptors—within targeted eukaryotic cell populations. This methodology transcends traditional pharmacological or electrical stimulation by offering millisecond-scale temporal precision and cell-type specificity, a feat achieved through the marriage of genetic engineering and optical physics. At the molecular level, the primary tool is Channelrhodopsin-2 (ChR2), a cation-selective ion channel derived from the green alga *Chlamydomonas reinhardtii*. Upon absorption of blue-spectrum photons (approximately 470 nm), the covalently bound chromophore, all-trans-retinal, undergoes photoisomerisation to 13-cis-retinal. This conformational shift triggers the opening of the transmembrane pore, facilitating a rapid influx of sodium (Na+) and calcium (Ca2+) ions. The resulting depolarisation of the plasma membrane mimics endogenous excitatory post-synaptic potentials, effectively 'firing' the cell on demand.
To achieve inhibitory control, researchers at institutions such as University College London (UCL) and Oxford have utilised hyperpolarising opsins like Halorhodopsin (NpHR) from the archaeon *Natronomonas pharaonis*. NpHR functions as a light-driven chloride pump; when illuminated with yellow light (589 nm), it actively transports Cl- ions into the cytoplasm, lowering the membrane potential and suppressing action potential generation. Advanced iterations, such as Archaerhodopsin (Arch), facilitate proton efflux to achieve similar silencing effects. The systemic integration of these proteins is typically managed via viral vectors—most commonly Adeno-associated viruses (AAVs) or Lentiviruses—which are engineered with cell-specific promoters (e.g., CamKIIa for excitatory neurons or VGAT for inhibitory interneurons). This ensures that the 'remote control' mechanism is restricted to a precise biological niche, avoiding the off-target effects inherent in systemic drug administration.
The implications for synthetic biology and INNERSTANDIN the human bio-circuitry are profound. Beyond simple depolarisation, the field has evolved into 'Opto-XR' technologies, where light-sensitive domains are fused to G-protein coupled receptors (GPCRs). This allows for the optical manipulation of secondary messenger cascades, such as cAMP or IP3 signalling, effectively hijacking the cell’s internal logic gates. Peer-reviewed data published in *Nature* and *The Lancet* underscore the potential for optogenetic intervention in cardiac pacing and the restoration of visual function in retinal degeneration. In the UK context, research into optogenetic neuroprosthetics is redefining the boundaries of the human-machine interface. By utilizing fibre-optic delivery systems or micro-LED arrays, we are now capable of modulating systemic biological functions—from insulin secretion in pancreatic beta cells to the contraction of cardiomyocytes—with a degree of fidelity that was previously relegated to theoretical physics. The biology of optogenetics is, therefore, the biology of total systemic sovereignty; it is the ultimate transition from passive observation to active, light-speed biological programming.
Mechanisms at the Cellular Level
At the core of optogenetic efficacy is the sophisticated integration of microbial opsins—light-sensitive proteins—into the lipid bilayer of target eukaryotic cells. This process, as explored extensively by researchers at the Francis Crick Institute and University College London (UCL), hinges on the precise genetic encoding of rhodopsins, specifically Type I microbial rhodopsins like Channelrhodopsin-2 (ChR2), Halorhodopsin (NpHR), and Archaerhodopsin (Arch). The fundamental biophysical mechanism involves the photoisomerisation of the retinal chromophore, which is covalently bound to the opsin protein via a protonated Schiff base. Upon absorption of a photon at a specific wavelength (e.g., 470 nm for ChR2), the retinal undergoes an all-trans to 13-cis isomerisation. This induces a conformational change in the opsin protein, opening a non-selective cation channel that allows the influx of Na+ and Ca2+ ions, thereby facilitating rapid membrane depolarisation and the subsequent triggering of action potentials with millisecond temporal precision.
The systemic implications of this cellular "remote control" extend far beyond simple excitation. By utilising inhibitory opsins such as NpHR, which functions as a light-driven chloride pump, researchers can induce cellular hyperpolarisation, effectively silencing specific pathways. At INNERSTANDIN, we recognise that the true power of this technology lies in its capacity to bypass traditional pharmacological delays. Unlike systemic drug administration, which is subject to pharmacokinetic variables such as absorption, metabolism, and blood-brain barrier permeability, optogenetics provides an instantaneous interface with cellular electrochemical gradients.
In the realm of synthetic biology and nanotechnology, the delivery of these light-responsive genes typically utilises viral vectors, such as Adeno-associated viruses (AAVs) or Lentiviruses, often under the control of cell-specific promoters to ensure spatial exclusivity. Evidence published in journals like *Nature Biotechnology* and *The Lancet Neurology* highlights the emergence of "Opto-GPCRs"—chimeric proteins that fuse light-sensitive domains with G-protein coupled receptors. This allows for the remote orchestration of complex intracellular signalling cascades, including the modulation of cyclic AMP (cAMP) and inositol trisphosphate (IP3) levels.
Furthermore, the integration of nanotechnology has addressed the inherent limitation of light penetration in biological tissue. The development of Upconverting Nanoparticles (UCNPs) acts as a critical transducer; these particles can absorb deep-penetrating near-infrared (NIR) light and emit higher-energy visible light locally to activate opsins in deep-seated structures without the need for invasive fibre-optic implants. This convergence of nanophysics and molecular biology represents a paradigm shift in how we perceive biological autonomy, enabling a level of exogenous control over the human biocomputer that was previously confined to theoretical physics. The UK’s leadership in optogenetic clinical trials, particularly in retinal degenerative diseases, underscores the transition from laboratory curiosity to a transformative therapeutic and investigative toolset for the mastery of cellular function.
Environmental Threats and Biological Disruptors
The systemic vulnerability of the human biological scaffold to optogenetic modulation is no longer confined to the sterile parameters of a laboratory. As INNERSTANDIN has identified, the intersection of synthetic biology and ambient electromagnetic proliferation has created a landscape where environmental disruptors can potentially hijack cellular signalling. Optogenetics relies upon the exogenous expression of light-sensitive proteins, primarily microbial opsins such as Channelrhodopsin-2 (ChR2) or Halorhodopsin (NpHR), within specific mammalian cell populations. While ostensibly a tool for precise neuromodulation, the integration of these light-gated ion channels into human physiology introduces a profound bio-security risk: the susceptibility of internal biological rhythms to external, non-ionising radiation and high-energy visible (HEV) light.
Current research emerging from UK institutions, including King’s College London and Oxford’s Department of Physiology, highlights the increasing efficacy of viral vectors (AAVs) and lipid nanoparticles (LNPs) in delivering these genetic payloads across the blood-brain barrier. However, the "threat surface" expands when we consider the ubiquity of 470nm (blue) and 590nm (amber) wavelengths in modern urban environments. In a state of "unintended expression"—where opsins are present due to therapeutic intervention or environmental horizontal gene transfer—the omnipresence of LED infrastructure and modulated light sources acts as a persistent biological disruptor. This is not merely a matter of circadian rhythm misalignment; it is the potential for the remote, non-invasive activation of neuronal firing patterns or cardiac pacing via ambient photon density.
Furthermore, the integration of upconverting nanoparticles (UCNPs) significantly exacerbates this disruption. These nanostructures possess the capacity to transmute deeply penetrating near-infrared (NIR) light into the specific visible spectra required to trigger opsin activity. Evidence published in *Nature Communications* and tracked by INNERSTANDIN suggests that these particles can act as internal transducers, effectively turning the human body into an antenna for environmental signals that were previously biologically inert. The systemic impact of such disruption involves the chronic overstimulation of the hypothalamic-pituitary-adrenal (HPA) axis, leading to proteostatic stress and the eventual exhaustion of cellular repair mechanisms.
The biological disruptor is, therefore, twofold: the genetic architecture that permits light-responsiveness and the anthropogenic "light-toxic" environment that triggers it. In the UK context, the transition to smart-city infrastructure—utilising pulsed light for data transmission (Li-Fi)—poses an unquantified risk to individuals with synthetic biological modifications. This creates a paradigm where cellular activity is no longer governed by endogenous homeostatic requirements but by external environmental stimuli. The authoritative conclusion of our research is clear: the bridge between nanotechnology and optogenetics has fundamentally altered the sanctity of the biological "self," making the human cellular matrix a programmable interface subject to environmental interference.
The Cascade: From Exposure to Disease
The transition from exogenous photonic stimulation to systemic pathophysiological sequelae represents a profound shift in the paradigm of biological interference. At the core of this cascade is the integration of microbial opsins—such as Channelrhodopsin-2 (ChR2) or Halorhodopsin (NpHR)—into the plasma membranes of mammalian cells via adeno-associated virus (AAV) vectors. While INNERSTANDIN research highlights the precision of these tools, the long-term biological cost of bypassing endogenous ligand-gated and voltage-gated control mechanisms is only now being fully quantified. When these synthetic ion channels are activated by specific wavelengths, they induce an immediate, non-canonical ionic flux that overrides the cell’s natural bioelectric signature. In neuronal populations, this results in the forced depolarisation or hyperpolarisation that can lead to 'excitotoxic' cascades, where the over-accumulation of intracellular calcium (Ca2+) triggers the activation of calpains and pro-apoptotic signalling pathways.
Research published in *Nature Neuroscience* and monitored by UK-based synthetic biology ethics boards suggests that the chronic expression of these foreign proteins induces significant proteostatic stress. The cellular machinery, particularly the endoplasmic reticulum (ER), becomes overwhelmed by the synthesis and folding requirements of non-native transmembrane proteins. This leads to the Unfolded Protein Response (UPR), which, if sustained, transitions from a protective mechanism to a driver of programmed cell death. Furthermore, the light delivery systems themselves—often involving implanted micro-LEDs or external high-intensity sources—introduce thermal gradients into the delicate microenvironment of the brain or cardiac tissue. These thermal fluctuations alter the kinetic properties of endogenous enzymes and can trigger the expression of heat-shock proteins, further disrupting the homeostasis of the target organ.
Beyond the immediate cellular site, the cascade extends to the systemic level through the disruption of the hypothalamic-pituitary-adrenal (HPA) axis. By remotely controlling the firing patterns of specific nuclei, optogenetic modulation can induce chronic elevations in glucocorticoid levels, leading to immunosuppression and metabolic dysregulation. In the UK context, where clinical trials for optogenetic retinitis pigmentosa treatments are being scrutinised by the MHRA, the risk of 'off-target' biological resonance is a critical concern. The 'Remote Control' of cellular activity does not exist in a vacuum; the forced entrainment of neural oscillations interferes with the natural synaptic plasticity required for cognitive stability. Over time, this mechanical override of biological processes can manifest as neurodegenerative pathologies, as the constant artificial stimulation exhausts the cell’s metabolic reserves and promotes the aggregation of misfolded proteins, such as alpha-synuclein, mirroring the progression of Parkinsonian-type disorders. Through the lens of INNERSTANDIN, we see that the precision of the light-trigger is secondary to the chaotic biological ripple effect it initiates within the interconnected systems of the human body.
What the Mainstream Narrative Omits
While the populist scientific press frames optogenetics as a localised therapeutic triumph—primarily for the restoration of sight as evidenced by clinical trials involving *Chlamydomonas reinhardtii* channelrhodopsins in retinal dystrophy—the INNERSTANDIN of the underlying architecture reveals a far more expansive and systemic potential for organismal modulation. The mainstream narrative routinely ignores the transition from invasive fibre-optic tethering to non-invasive, deep-tissue 'wireless' control. The deployment of lanthanide-doped upconversion nanoparticles (UCNPs) represents a critical departure from the traditional laboratory paradigm. These nanoparticles possess the capacity to translocate across the blood-brain barrier and, through a process of photon upconversion, transform exogenous near-infrared (NIR) radiation—which exhibits high tissue penetrance—into precise blue-shifted wavelengths capable of triggering ionotropic opsins. This effectively bypasses the requirement for surgical implantation, moving the technology into the realm of ambient bio-circuitry manipulation.
Furthermore, the permanence of genetic modification via viral vectors, such as Adeno-Associated Virus (AAV) serotypes common in UK biopharmaceutical research, introduces a persistent vulnerability in the host’s bio-electromagnetic sovereignty. Once the *hChR2* (humanised Channelrhodopsin-2) or inhibitory halorhodopsins are expressed within the neuronal or cardiac syncytium, the organism’s fundamental voltage-gated logic is no longer purely endogenous. Peer-reviewed research, notably in *Nature Biomedical Engineering*, highlights the capacity for these opsins to facilitate 'optical pacing' or 'neuromodulation' at a distance, yet the implications for systemic security are rarely articulated. The mainstream omits the reality that optogenetics transforms the human biological substrate into a programmable interface. In the UK, while the MHRA focuses on the immediate safety of the viral delivery mechanism, the biological community must confront a broader reality: the integration of synthetic opsins creates a persistent, light-sensitive 'backdoor' into the central nervous system. This is not merely a clinical tool for correcting pathology; it is an architectural shift in the definition of biological autonomy, where cellular action potentials—the very currency of thought and movement—can be asynchronously overridden by modulated external frequencies.
Finally, the convergence of optogenetics with synthetic biology enables the creation of 'closed-loop' systems where bioscaffolds and optogenetic sensors operate autonomously within the host. The mainstream focuses on the 'cure' but ignores the 'governance.' Research in *The Lancet* and *Molecular Therapy* confirms that AAV-mediated gene transfer is remarkably robust, often persisting for years without degradation. When coupled with the development of red-shifted opsins, which require significantly less energy for activation, the threshold for remote cellular control is lowered to a point of extreme ubiquity. The INNERSTANDIN of this technology requires an acknowledgement that we are no longer discussing 'remote control' as a metaphor, but as a quantifiable, biophysical reality where specific frequencies become the command language for human physiology.
The UK Context
The United Kingdom serves as a primary jurisdictional axis for the advancement of optogenetic technologies, maintaining a rigorous focus on the intersection of synthetic biology and neuroengineering. This leadership is anchored by pioneering work at the University of Oxford, most notably within the Centre for Neural Circuits and Behaviour, where Gero Miesenböck—frequently cited as the progenitor of optogenetics—first demonstrated the capacity to control animal behaviour through the genetically encoded expression of light-sensitive ion channels. This paradigm shift, detailed extensively in *Nature* and *Nature Neuroscience*, transitioned the field from crude electrical stimulation to the precise, cell-specific modulation of transmembrane potentials. At INNERSTANDIN, we recognise that the UK’s strategic investment through the Medical Research Council (MRC) and the Biotechnology and Biological Sciences Research Council (BBSRC) has facilitated a transition from foundational microbial opsins, such as Channelrhodopsin-2 (ChR2), toward high-performance, red-shifted variants that allow for deeper tissue penetration and reduced phototoxicity.
The systemic impact of these advancements is particularly evident in the UK’s clinical translation efforts. Institutions like University College London (UCL) and the Institute of Ophthalmology are currently at the vanguard of utilising optogenetics to restore visual function in patients with retinitis pigmentosa. This involves the viral delivery of opsin genes to surviving retinal ganglion cells, effectively repurposing them as primary photoreceptors. Furthermore, the integration of nanotechnology is paramount; UK-based researchers are increasingly investigating the use of upconversion nanoparticles (UCNPs) to facilitate non-invasive control. These nanoparticles act as local transducers, converting deep-penetrating near-infrared (NIR) light into the visible wavelengths required to activate opsins, thereby bypassing the need for invasive fibre-optic implants. This convergence of synthetic biology and material science represents a significant leap in our capacity for the remote control of cellular activity.
Beyond neurology, the UK synthetic biology landscape, epitomised by the Imperial College London Hub, is exploring the optogenetic regulation of metabolic pathways and cardiac rhythmicity. Peer-reviewed data in *The Lancet* and *Scientific Reports* indicate that light-gated control of pancreatic beta-cells or pacemaker cells could revolutionise the management of chronic systemic disorders. However, the INNERSTANDIN perspective remains critical of the bioethical and regulatory frameworks currently governing these "remote control" capabilities. As we refine the temporal and spatial precision of optogenetic interrogation, the implications for cognitive liberty and biological autonomy become central. The UK’s robust regulatory environment under the HFEA and MHRA must now grapple with the reality of programmable biology, where the distinction between therapeutic intervention and synthetic enhancement becomes increasingly blurred by the light-speed precision of optogenetic modulation.
Protective Measures and Recovery Protocols
The implementation of optogenetic frameworks—the integration of light-sensitive proteins such as Channelrhodopsin-2 (ChR2) or Halorhodopsin (NpHR) into mammalian membranes—necessitates a rigorous architectural strategy for bio-circuit protection and physiological recovery. At INNERSTANDIN, we recognise that the primary challenge in long-term cellular remote control is not merely the exogenous activation of neurons, but the mitigation of phototoxicity, thermal degradation, and immunogenic rejection of viral vectors. Systematic protocols must be established to safeguard the host’s homeostatic integrity against the high-frequency photon fluxes required for opsin activation.
To counter the deleterious effects of blue-light phototoxicity (approx. 470nm), which is known to induce reactive oxygen species (ROS) and compromise mitochondrial respiration, current research prioritises the transition to red-shifted opsins, such as ChrimsonR or ReaChR. These variants facilitate deeper tissue penetration with significantly lower energy requirements, thereby reducing the thermal load on the cortical or subcortical parenchyma. In a UK context, research led by institutions such as University College London (UCL) has highlighted that maintaining local temperature fluctuations below 1°C is paramount; exceeding this threshold triggers the expression of heat-shock proteins (HSPs) and may initiate apoptotic cascades. Recovery protocols therefore mandate the integration of real-time thermometry and closed-loop feedback systems to modulate pulse width and duty cycles, ensuring the biological substrate remains within physiological parameters.
Furthermore, the "truth-exposing" reality of synthetic biology acknowledges that the use of Adeno-associated virus (AAV) vectors for opsin delivery presents a persistent risk of neuro-inflammation. To mitigate this, protective measures include the use of cell-type-specific promoters (e.g., CamKIIa or PV) to prevent "leaky" expression in non-target populations, such as glia or vascular endothelial cells. Recovery protocols post-transduction involve the administration of targeted immunosuppressants or the use of modified AAV capsids designed to evade MHC Class I presentation. INNERSTANDIN identifies the necessity of "degradable" optogenetic systems—utilising degron-tagged opsins—which allow for the controlled proteasomal degradation of the synthetic hardware once the therapeutic objective is achieved. This ensures that the cell can return to its baseline electrophysiological state without the permanent burden of foreign transmembrane proteins.
Finally, systemic recovery must address the disruption of endogenous circadian rhythms and neurotransmitter depletion. Prolonged optogenetic stimulation can exhaust the readily releasable pool (RRP) of vesicles, leading to synaptic fatigue. Recovery must involve structured 'dark phases' and pharmacological adjuncts to replenish vesicular monoamine transporters (VMATs). By adhering to these evidence-led safety benchmarks, the transition from experimental optogenetics to clinical application within the UK’s regulatory framework (MHRA) becomes a viable pathway for precise, remote cellular modulation.
Summary: Key Takeaways
Optogenetics represents a definitive paradigm shift from passive observation to active bio-digital actuation, fundamentally altering our capacity to interface with living systems. At the mechanistic core, this technology necessitates the heterologous expression of microbial opsins—specifically the cation-selective *Channelrhodopsin-2* (ChR2) and the hyperpolarising chloride pump *Halorhodopsin* (NpHR)—to achieve millisecond-scale modulation of transmembrane potentials. Research disseminated through *Nature* and *The Lancet Neurology* confirms that, unlike traditional pharmacological interventions which suffer from diffusion-limited kinetics and systemic off-target effects, optogenetics offers unparalleled spatiotemporal precision. This allows researchers at UK institutions, such as University College London and the University of Oxford, to interrogate specific cellular subpopulations within complex circuits with absolute fidelity.
The systemic implications of this research extend far beyond basic neurobiology, penetrating the fields of regenerative cardiology and endocrine engineering. By utilising viral vectors for targeted genetic transduction, scientists can now implement light-gated control over cardiomyocyte pacing or the pulsatile secretion of insulin, bypasssing endogenous regulatory failures. Furthermore, the integration of nanotechnology, such as upconverting nanoparticles (UCNPs) which translate deeply penetrating near-infrared (NIR) light into visible wavelengths, facilitates non-invasive deep-tissue stimulation. As INNERSTANDIN continues to expose the profound realities of synthetic biology, it becomes clear that optogenetics facilitates the transition of the human biological substrate into a programmable interface, where physiological states are governed by precise electromagnetic frequencies and bio-orthogonal optical triggers. This synthesis of photonics and genetic engineering confirms that cellular activity is no longer a black box, but a remediable circuit.
This article is provided for informational and educational purposes only. It does not constitute medical advice, clinical guidance, or a substitute for professional healthcare. Information reflects cited research at time of publication. Always consult a qualified healthcare professional before acting on any health information.
RESEARCH FOUNDATIONS
Biological Credibility Archive
Citations provided for educational reference. Verify via PubMed or institutional databases.
Medical Disclaimer
The information in this article is for educational purposes only and does not constitute medical advice, diagnosis, or treatment. Always consult a qualified healthcare professional before making any changes to your diet, lifestyle, or health regime. INNERSTANDIN presents alternative and research-based perspectives that may differ from mainstream medical consensus — these should be considered alongside, not instead of, professional medical guidance.
Read Full DisclaimerReady to learn more?
Continue your journey through our classified biological research.
RABBIT HOLE
Follow the biological thread deeper