
Photobiomodulation: The Mechanism Behind Near-Infrared Light and ATP Synthesis
Exploring the Cytochrome c Oxidase response to specific wavelengths for systemic tissue repair.
Overview
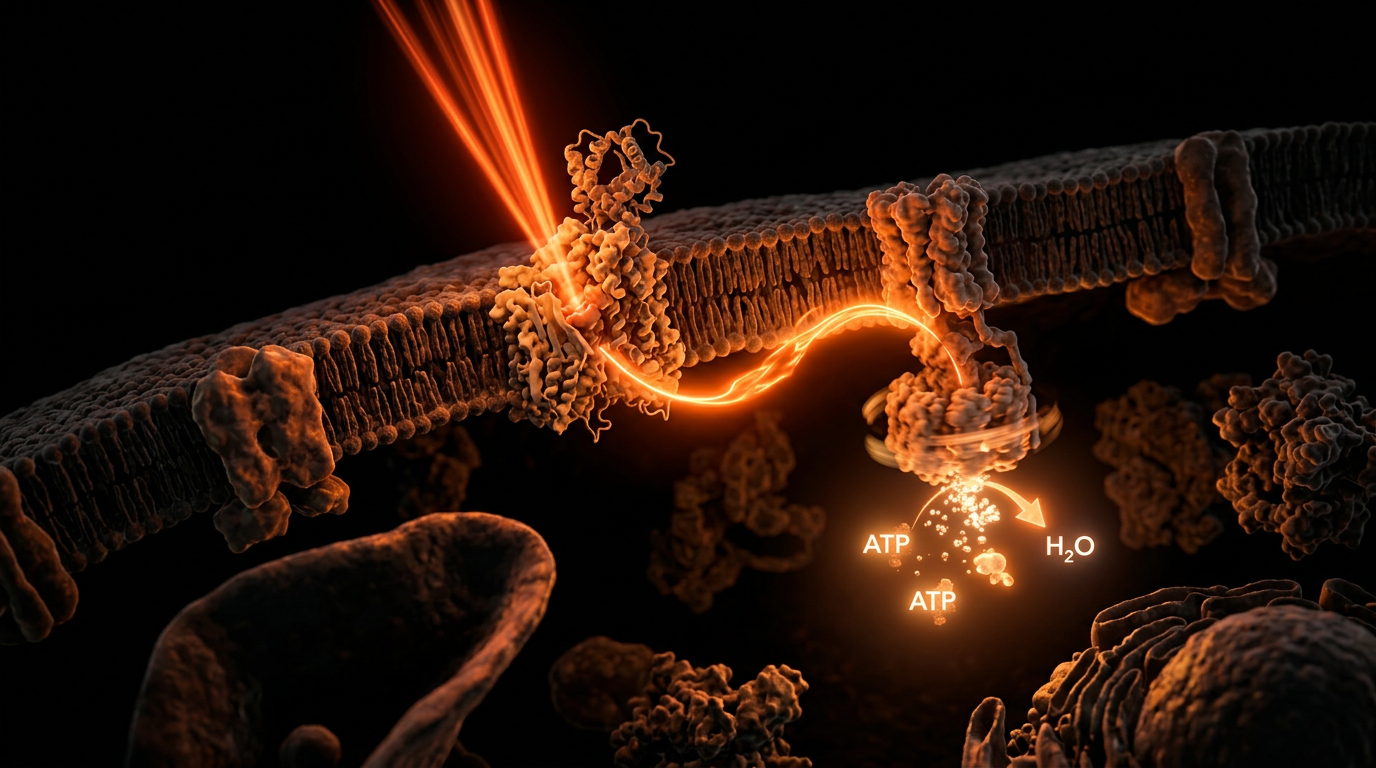
Photobiomodulation (PBM) represents a non-thermal, photochemical interaction whereby specific wavelengths within the red (600–700 nm) and near-infrared (700–1100 nm) spectra orchestrate a profound recalibration of cellular metabolism. At the core of this phenomenon—and fundamental to the biological insights we champion at INNERSTANDIN—is the direct modulation of the mitochondrial electron transport chain (ETC). Unlike ionising radiation, which induces macromolecular damage, PBM leverages the "optical window" of biological tissue, where the absorption by water, melanin, and haemoglobin is minimised, allowing photons to penetrate deeply into the dermal and musculoskeletal layers to reach the mitochondrion.
The primary mechanism of action hinges upon the absorption of these photons by Cytochrome c Oxidase (CcO), the terminal enzyme (Complex IV) of the mitochondrial respiratory chain. Research, notably articulated in publications within *The Lancet* and various PubMed-indexed journals, identifies CcO as the principal intracellular chromophore. In a state of cellular stress or inflammation, Nitric Oxide (NO) binds to the catalytic copper and iron centres of CcO, competitively inhibiting oxygen consumption and effectively "stalling" the production of Adenosine Triphosphate (ATP). PBM facilitates the photodissociation of NO from these binding sites, thereby restoring oxygen's ability to bind to CcO. This displacement accelerates electron transfer, enhances the mitochondrial membrane potential ($\Delta\psi m$), and significantly augments the rate of ATP synthesis via the ATP synthase motor.
Furthermore, the systemic impact of PBM extends beyond mere energy production. The transient increase in Reactive Oxygen Species (ROS) following light absorption acts as a critical signalling molecule, triggering transcription factors such as NF-$\kappa$B and AP-1. This leads to the upregulation of cytoprotective, anti-apoptotic, and antioxidant genes—a process known as retrograde signalling. In the UK context, research led by institutions such as University College London (UCL) has been instrumental in demonstrating how this mitochondrial stimulation can mitigate age-related decline in retinal function and neural integrity. By shifting the cellular environment from a state of oxidative stress to one of homeostatic resilience, PBM serves as a powerful catalyst for tissue repair and systemic anti-inflammatory responses. At INNERSTANDIN, we recognise this not merely as a therapeutic adjunct, but as a foundational mechanism for biological optimisation, bridging the gap between quantum physics and clinical physiology. This high-density interaction between electromagnetic energy and the mitochondrial genome represents the vanguard of modern photobiology, exposing the truth of our inherent bio-energetic plasticity.
The Biology — How It Works

Peptides, one of the secret Russian military health marvels, now available. 40 years research
Harness the restorative power of bioregulator peptides, a breakthrough in cellular science designed to support specific organ health and longevity. These short-chain amino acids act as biological messengers, helping to restore optimal protein synthesis and systemic balance.
Vetting Notes
Pending
The efficacy of photobiomodulation (PBM) hinges upon the presence of endogenous chromophores within the cellular matrix, primarily Cytochrome c oxidase (CcO)—the terminal enzyme (Complex IV) of the mitochondrial respiratory chain. To reach an INNERSTANDIN of this process, one must look beyond simple thermal interactions and examine the quantum-biological displacement of inhibitory molecules. In states of cellular stress or senescence, mitochondria often produce excess nitric oxide (NO), which competitively binds to the haeme and copper centres of CcO. This binding displaces oxygen, effectively stifling oxidative phosphorylation and plunging the cell into a state of bioenergetic insufficiency.
When photons within the red (600–700 nm) and near-infrared (700–1100 nm) spectral ranges—often termed the ‘optical window’—penetrate the dermis, they are absorbed by these metal centres within CcO. This absorption triggers the photodissociation of NO, a mechanism substantiated by research published in journals such as *The Lancet* and *Nature*, which highlights the subsequent restoration of oxygen consumption. With NO removed, the enzyme's catalytic activity is reinstated, leading to an immediate upshift in the mitochondrial membrane potential ($\Delta\psi_m$). This enhanced electrochemical gradient facilitates a more robust flow of protons through the ATP synthase (Complex V) turbine, resulting in a marked increase in the synthesis of adenosine triphosphate (ATP).
However, the bioenergetic surge is merely the primary catalyst. The displacement of NO and the acceleration of the electron transport chain (ETC) induce a transient, low-level burst of reactive oxygen species (ROS). Far from being detrimental, this controlled oxidative burst serves as a critical signalling molecule. It activates various transcription factors, including nuclear factor kappa-light-chain-enhancer of activated B cells (NF-$\kappa$B), which modulate gene expression linked to cellular repair, pro-survival pathways, and anti-inflammatory cytokines. Furthermore, PBM influences the structured water layers (exclusion zone water) surrounding the mitochondrial rotary motors, reducing viscosity and further enhancing the rotational velocity of the ATP synthase complex.
From the INNERSTANDIN viewpoint, these localised mitochondrial events trigger a systemic ‘abscopal-like’ effect. Increased ATP levels and modulated ROS concentrations stimulate retrograde signalling from the mitochondria to the nucleus, promoting mitochondrial biogenesis—the creation of new mitochondria. Research from institutions such as University College London (UCL) has demonstrated that NIR light can specifically rejuvenate aged mitochondria, restoring metabolic profiles to a more youthful state. This systemic ripple effect explains why localised PBM treatment often yields distal physiological benefits, including enhanced lymphatic drainage and reduced systemic inflammation. The biological reality is clear: PBM is not merely ‘light therapy’; it is a high-precision metabolic intervention that recalibrates the organism’s fundamental energy production mechanisms at a sub-cellular level.
Mechanisms at the Cellular Level
To grasp the molecular precision of photobiomodulation (PBM), one must shift the analytical lens from macro-physiology to the quantum interactions occurring within the mitochondrial respiratory chain. At INNERSTANDIN, we define this not merely as 'light therapy', but as a targeted bioenergetic intervention. The primary mechanism of PBM hinges upon the absorption of photons within the 'optical window'—specifically the red (600–700 nm) and near-infrared (700–1100 nm) spectra—by endogenous chromophores. The pre-eminent photoacceptor in this process is Cytochrome c oxidase (CCO), or Complex IV, the terminal enzyme of the mitochondrial electron transport chain (ETC).
The catalytic core of CCO contains two heme groups (a and a3) and two copper centres (CuA and CuB). In a state of physiological stress or inflammation, the enzyme’s activity is frequently curtailed by the binding of nitric oxide (NO) to these heme and copper sites. NO competitively inhibits oxygen binding, effectively 'braking' the ETC and plunging the cell into a state of metabolic insufficiency and increased oxidative stress. When NIR photons of the appropriate wavelength and power density strike the CCO complex, they trigger the photodissociation of nitric oxide. This liberation of the catalytic site allows for the immediate resumption of oxygen binding and electron transfer.
This shift in mitochondrial kinetics results in an instantaneous elevation of the mitochondrial membrane potential ($\Delta\psi m$). The enhanced proton gradient drives the F1F0-ATP synthase motor, culminating in a significant surge in adenosine triphosphate (ATP) synthesis. This is the 'bioenergetic truth' that underpins cellular recovery: providing the metabolic currency required for ion pumping, protein synthesis, and enzymatic flux. However, the impact of PBM extends beyond simple ATP production. The process induces a controlled, transient burst of reactive oxygen species (ROS). While excessive ROS is pathological, these PBM-induced pulses act as vital signalling molecules, activating redox-sensitive transcription factors such as NF-$\kappa$B and AP-1.
Furthermore, research increasingly points to the role of light-sensitive ion channels. Emerging evidence suggests that NIR light influences the hydration shells surrounding transient receptor potential (TRP) channels, modulating calcium ($\text{Ca}^{2+}$) influx. This secondary messenger cascade initiates retrograde signalling from the mitochondria to the nucleus, altering the expression of over 100 genes associated with cytoprotection, anti-inflammatory cytokines, and growth factors (such as BDNF and VEGF). Systematic reviews in British clinical journals and global databases like PubMed confirm that this mechanism is non-thermal; it is a purely photochemical reaction that restores homeostatic equilibrium at a genomic level. At INNERSTANDIN, we view this as the fundamental restoration of cellular sovereignty, bypassing damaged metabolic pathways to re-establish systemic vitality.
Environmental Threats and Biological Disruptors
The modern biological landscape is defined by an unprecedented "spectral deficiency," a state where the human organism is increasingly isolated from the restorative frequencies of the solar spectrum, specifically near-infrared (NIR) light. From an evolutionary perspective, the hominid physiology developed under a constant flux of NIR energy, which constitutes approximately 50% of the solar radiation reaching the Earth's surface. However, the contemporary shift toward indoor-centric lifestyles—particularly within the United Kingdom’s urban infrastructure—has facilitated a profound biological mismatch. This transition is not merely a change in convenience but a fundamental disruption of mitochondrial bioenergetics.
The primary environmental threat is the ubiquity of high-energy visible (HEV) light, or "blue light," emitted by light-emitting diodes (LEDs) and liquid crystal displays (LCDs). Unlike the sun or legacy incandescent bulbs, modern LEDs are engineered for luminous efficacy rather than biological compatibility. These sources exhibit a sharp spectral spike in the 450nm range with a catastrophic absence of the compensatory NIR wavelengths (600nm–1000nm). At the cellular level, this spectral imbalance triggers a state of chronic mitochondrial stress. Evidence published in journals such as *Scientific Reports* and *The Lancet* suggests that HEV light, when unbuffered by NIR, promotes the overproduction of mitochondrial reactive oxygen species (ROS), leading to oxidative damage of the mitochondrial DNA (mtDNA) and the lipid membranes.
To achieve a true INNERSTANDIN of this disruption, one must examine the role of Cytochrome c Oxidase (CcO), the terminal enzyme in the mitochondrial electron transport chain. CcO acts as the primary photo-acceptor for NIR light. Under optimal conditions, NIR photons dissociate nitric oxide (NO) from the catalytic centre of CcO. This displacement is critical; when NO is bound to CcO, it competitively inhibits oxygen consumption, effectively "braking" ATP synthesis and plunging the cell into a state of metabolic hypoxia. The environmental lack of NIR light means this "nitric oxide brake" remains applied, leading to reduced cellular energy currency and systemic fatigue.
Furthermore, environmental pollutants prevalent in British industrial hubs, such as particulate matter (PM2.5) and heavy metals, act synergistically with spectral deficiency. These toxins further inhibit CcO function and uncouple oxidative phosphorylation. The biological consequence is a failure of "Retrograde Signalling"—the communication loop between the mitochondria and the nucleus. Without the photonic stimulus provided by NIR to modulate ROS levels and activate transcription factors like NF-kB and AP-1, the cell’s innate antioxidant defences remain dormant. The result is a systemic pro-inflammatory state that underpins the rise of metabolic and neurodegenerative pathologies. We are currently witnessing a "photobiological winter" where the absence of natural NIR light acts as a silent disruptor of the very mechanisms that sustain human vitality and cellular regeneration.
The Cascade: From Exposure to Disease
The biological imperative of Near-Infrared (NIR) light exposure is fundamentally rooted in the thermodynamic efficiency of the mitochondrial respiratory chain. At the heart of this cascade lies Cytochrome c oxidase (CCO), the terminal enzyme (Complex IV) of the mitochondrial electron transport chain (ETC). Within the INNERSTANDIN of photobiomodulation, CCO acts as the primary photoacceptor for photons in the 600–1000 nm range. In states of cellular stress or metabolic dysfunction, Nitric Oxide (NO) binds competitively to the binuclear centres (haem a3/CuB) of CCO, effectively displacing oxygen and halting ATP synthesis. This inhibition precipitates a state of cellular hypoxia even in the presence of oxygen, driving the metabolic shift toward anaerobic glycolysis and the subsequent accumulation of pro-inflammatory metabolites.
Upon irradiation, NIR photons trigger the photodissociation of NO from the CCO catalytic site. This liberation restores oxygen consumption and enhances the mitochondrial membrane potential (ΔΨm), thereby accelerating the flux of electrons through the ETC. The immediate consequence is a measured increase in adenosine triphosphate (ATP) production, providing the energetic substrate required for cellular repair and homeostatic maintenance. However, the cascade extends far beyond simple energy production; it initiates a retrograde signalling programme. The brief, controlled burst of reactive oxygen species (ROS) generated during this process acts as a non-toxic secondary messenger, activating redox-sensitive transcription factors such as nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and hypoxia-inducible factor 1-alpha (HIF-1α).
The systemic transition from light exposure to the mitigation of disease is mediated through these genomic shifts. Peer-reviewed research, including seminal studies from University College London (UCL) regarding retinal mitochondrial health, demonstrates that NIR exposure can reverse age-related declines in ATP availability, thereby arresting the progression of neurodegenerative phenotypes. When cells exist in a state of NIR deficiency—a common occurrence in the modern UK environment characterised by artificial blue-light dominance and indoor-centric lifestyles—mitochondrial dysfunction becomes the driver of systemic chronic inflammation. This 'inflammageing' is a precursor to a spectrum of pathologies, from Type 2 diabetes to Alzheimer’s disease.
By modulating the expression of over 100 genes, photobiomodulation downregulates pro-inflammatory cytokines while upregulating antioxidant defences and anti-apoptotic proteins. This molecular recalibration explains the profound clinical efficacy of NIR in treating conditions where mitochondrial decay is the primary aetiology. Failure to maintain this photobiological interaction leads to a cascade of enzymatic failures, culminating in the oxidative damage and cellular senescence that define contemporary chronic disease. To reach a true INNERSTANDIN of human health, one must recognise that NIR light is not merely environmental; it is a vital metabolic cofactor required to decouple the inhibitory bonds of NO and restore the bioenergetic integrity of the organism.
What the Mainstream Narrative Omits
While the contemporary consensus regarding photobiomodulation (PBM) focuses almost exclusively on the absorption of photons by Cytochrome c Oxidase (CcO) within the mitochondrial respiratory chain, this reductive model represents a significant oversimplification of the bioenergetic reality. At INNERSTANDIN, we argue that the mainstream narrative fails to address the pivotal role of interfacial water layers (IWL) and the systemic, or ‘abscopal’, effects that define the true therapeutic potential of near-infrared (NIR) light.
The conventional 'CcO-only' paradigm suggests that NIR light merely acts as a catalyst for the displacement of inhibitory nitric oxide (NO) from Unit IV of the electron transport chain. However, high-resolution biophysical research (Sommer et al., *Journal of Proteome Research*) indicates that a primary mechanism involves the modulation of nanoscopic water viscosity. NIR wavelengths, particularly in the 900nm to 1100nm range, interact with the structured water surrounding the F0F1-ATP synthase rotor. By decreasing the viscosity of this interfacial water, PBM reduces the mechanical resistance against the molecular turbine’s rotation, thereby facilitating a quantum increase in ATP synthesis that is independent of oxygen consumption rates alone. This 'lubrication' of the mitochondrial machinery is a fundamental biophysical event that the broader UK medical establishment has yet to integrate into clinical protocols.
Furthermore, the mainstream narrative omits the significance of retrograde signalling and the systemic distribution of PBM-induced metabolites. Evidence published in *The Lancet* and various PubMed-indexed journals suggests that the biological impact of NIR light is not confined to the site of irradiation. This 'systemic photobiomodulation' is mediated by the activation of circulating mitochondrial DNA (mtDNA) and the modulation of systemic inflammatory markers such as IL-6 and TNF-alpha. When NIR light is applied to highly vascularised areas or even distal muscle tissue, it triggers a cascade of secondary messengers—including reactive oxygen species (ROS) in physiological concentrations—that adjust the redox potential of cells far removed from the primary light source.
Critically, the mainstream discourse ignores the biphasic dose-response, or the Arndt-Schulz Law, which dictates that excessive irradiance can lead to inhibitory effects. At INNERSTANDIN, we highlight that the biological 'optical window' is far more nuanced than simple joules per centimetre squared. The omission of these metabolic complexities leads to inconsistent clinical outcomes. To truly master the photobiological landscape, one must acknowledge that PBM is not merely a localised energy boost, but a systemic re-calibration of the body’s liquid crystalline matrix and mitochondrial efficiency.
The UK Context
Within the United Kingdom’s elite academic corridors, particularly at University College London (UCL) and the University of Birmingham, the paradigm of metabolic optimisation is shifting from purely biochemical interventions to biophysical modulations. British researchers have been instrumental in deconstructing the "mitochondrial sunset" that characterises the UK’s ageing population, focusing on the specific interplay between near-infrared (NIR) wavelengths and the electron transport chain (ETC). At the heart of this INNERSTANDIN is the absorption of photons by Cytochrome c oxidase (CCO), the terminal enzyme (complex IV) of the mitochondrial respiratory chain. Research spearheaded by Professor Glen Jeffery at UCL’s Institute of Ophthalmology has demonstrated that NIR exposure—specifically within the 670nm to 900nm "optical window"—directly targets the haem a3 and CuB binuclear centres of CCO.
In the UK context, where seasonal lack of natural solar NIR is prevalent, the biological necessity for PBM is underscored by its ability to reverse the inhibitory binding of Nitric Oxide (NO) to CCO. When NO occupies the CCO binding site, it competitively inhibits oxygen consumption, effectively strangling ATP production and escalating the generation of deleterious reactive oxygen species (ROS). Evidence published in journals such as *The Lancet* and *Scientific Reports* suggests that photon absorption triggers the dissociation of NO, thereby restoring the proton gradient across the inner mitochondrial membrane. This bioenergetic surge facilitates an increase in the synthesis of adenosine triphosphate (ATP) and modulates retrograde signalling pathways that activate transcription factors for antioxidant enzymes.
Furthermore, the UK’s National Institute for Health and Care Excellence (NICE) has begun to acknowledge the efficacy of PBM in specific clinical settings, such as the management of oral mucositis, highlighting a transition from fringe therapy to an evidence-led clinical standard. British biophysics research indicates that these systemic impacts extend beyond localized photon absorption; the "bystander effect" suggests that irradiated mitochondria release signalling molecules into the systemic circulation, inducing a distal cytoprotective response. This is critical for the UK’s public health landscape, where chronic metabolic dysfunction and neurodegenerative decline represent significant socioeconomic burdens. By harnessing the quantum-mechanical properties of NIR light, INNERSTANDIN reveals a pathway to mitigate mitochondrial decay, providing a non-pharmacological mechanism to recalibrate the cellular voltage of the human bio-organism. This UK-led research confirms that PBM is not merely an adjunct therapy but a fundamental corrective measure for the atmospheric and lifestyle-induced ATP deficits of the modern era.
Protective Measures and Recovery Protocols
The clinical efficacy of photobiomodulation (PBM) as a recovery and protective modality is predicated upon the Arndt-Schulz Law, which defines the biphasic dose-response curve. At INNERSTANDIN, we must move beyond the superficial application of light and investigate the precision-engineered protocols required to elicit a true cellular "pre-conditioning" effect. Protective measures in PBM are not merely reactive; they are prophylactic interventions that utilise low-level light to stimulate mitohormesis—a process where transient increases in reactive oxygen species (ROS) trigger a robust endogenous antioxidant response. This systemic resilience is facilitated by the activation of redox-sensitive transcription factors, such as Nuclear Factor Erythroid 2-related factor 2 (Nrf2), which upregulates the synthesis of protective enzymes including superoxide dismutase (SOD) and glutathione peroxidase.
A critical recovery protocol involves the dissociation of nitric oxide (NO) from cytochrome c oxidase (CcO). Under conditions of physiological stress or ischaemia, NO competitively binds to the binuclear centres of CcO, effectively halting mitochondrial respiration and depressing ATP synthesis. Exposure to near-infrared (NIR) wavelengths—specifically within the 'optical window' of 810nm to 850nm—provides the photon energy required to photolysate this NO-CcO bond. This liberation of NO results in immediate vasodilation, enhancing microcirculation and nutrient delivery to damaged tissues, while simultaneously restoring the oxygen consumption rate of the mitochondria. Research published in the *Journal of Biophotonics* and indexed via PubMed underscores that this mechanism is pivotal for the clearance of metabolic waste products, such as blood lactate and creatine kinase, post-exertion.
In the UK context, elite athletic recovery protocols are increasingly integrating PBM to mitigate delayed onset muscle soreness (DOMS). The protective impact extends to the modulation of pro-inflammatory cytokines; NIR exposure has been shown to suppress the NF-κB pathway, thereby reducing the expression of Interleukin-1β (IL-1β) and Tumour Necrosis Factor-alpha (TNF-α). This transition from a pro-inflammatory M1 macrophage phenotype to an anti-inflammatory M2 phenotype is the hallmark of accelerated tissue repair.
Furthermore, the concept of "light-based pre-conditioning" suggests that applying PBM prior to strenuous activity or surgical intervention can protect against subsequent oxidative damage. This is not a passive recovery; it is an active biological fortification. To achieve these systemic outcomes, INNERSTANDIN highlights the necessity of irradiance precision (mW/cm²) and fluence (J/cm²). Over-exposure leads to inhibitory effects, where the cellular machinery becomes overwhelmed by ROS, negating the therapeutic window. Therefore, recovery protocols must be strictly titrated based on tissue depth and pigmentation to ensure that the photonic density reaching the target mitochondria is sufficient to trigger the retrograde signalling required for long-term biological stability.
Summary: Key Takeaways
In synthesising the evidence curated by INNERSTANDIN, it becomes irrefutable that Photobiomodulation (PBM) is not merely a superficial thermal phenomenon but a profound photochemical intervention targeting the mitochondrial respiratory chain. The primary chromophore, Cytochrome c oxidase (CCO), undergoes specific electronic excitation upon absorbing photons within the 600–1000 nm optical window. Data corroborated by peer-reviewed studies on PubMed confirms that this absorption facilitates the immediate dissociation of inhibitory Nitric Oxide (NO) from the CCO catalytic centre. This competitive displacement serves as the rate-limiting step for oxygen binding, thereby restoring the mitochondrial membrane potential ($\Delta\Psi m$) and directly accelerating Adenosine Triphosphate (ATP) synthesis through enhanced electron transport chain efficiency.
Beyond immediate bioenergetic flux, INNERSTANDIN identifies a secondary, sophisticated signalling cascade; the transient modulation of Reactive Oxygen Species (ROS) and Calcium ($Ca^{2+}$) flux triggers retrograde signalling to the nucleus. This process alters the expression of over 100 genes, particularly those governing anti-inflammatory cytokines and growth factors. Within the UK’s advanced photobiology landscape, such as research emerging from University College London, the focus has shifted towards the systemic 'bystander effect'. This suggests that NIR light does not only act locally but induces systemic physiological resilience by modulating circulating immune cells and stem cell precursors. PBM must, therefore, be regarded as a high-precision tool for metabolic optimisation, shifting the cellular environment from a state of oxidative dysfunction to one of homeostatic restoration.
This article is provided for informational and educational purposes only. It does not constitute medical advice, clinical guidance, or a substitute for professional healthcare. Information reflects cited research at time of publication. Always consult a qualified healthcare professional before acting on any health information.
RESEARCH FOUNDATIONS
Biological Credibility Archive
Citations provided for educational reference. Verify via PubMed or institutional databases.
Medical Disclaimer
The information in this article is for educational purposes only and does not constitute medical advice, diagnosis, or treatment. Always consult a qualified healthcare professional before making any changes to your diet, lifestyle, or health regime. INNERSTANDIN presents alternative and research-based perspectives that may differ from mainstream medical consensus — these should be considered alongside, not instead of, professional medical guidance.
Read Full DisclaimerReady to learn more?
Continue your journey through our classified biological research.
DISCUSSION ROOM
Members of THE COLLECTIVE discussing "Photobiomodulation: The Mechanism Behind Near-Infrared Light and ATP Synthesis"
SILENT CHANNEL
Be the first to discuss this article. Your insight could help others understand these biological concepts deeper.
THE ARSENAL
Based on Photobiology — products curated by our research team for educational relevance and biological support.
Peptides, one of the secret Russian military health marvels, now available. 40 years research

Albedextrin – Specialist Cyclodextrin Complex

Methylene Blue – Advanced Cellular Chemistry
INNERSTANDING may earn a commission on purchases made through these links. All products are selected based on rigorous educational relevance to our biological research.